
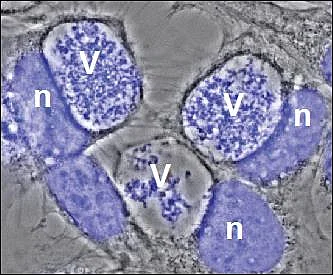
V= Vacuole filled with bacteria
n = host cell nucleus
Research
The main interest of our group is to understand host-pathogen interactions on the molecular level. We are studying molecular and cellular events that enable microbial pathogens to evade host defense mechanisms. In particular, we are interested in how obligate intracellular pathogens modulate host cell apoptosis pathways, signal transduction and vesicular trafficking. We are using the Coxiella burnetii as a model system. C. burnetii is the causative agent of human Q fever, a worldwide zoonotic disease. These small Gram-negative bacteria are classified in the y subdivision of proteobacteria. C. burnetii are typically transmitted to humans by inhalation of infectious material transmitted from domestic livestock. Q fever is often an asymptomatic or mild flu-like illness, but can develop into an atypical pneumonia or hepatitis. Furthermore, the infection can also result in chronic Q fever which is typically characterized by bacterial endocarditis and is potentially fatal.
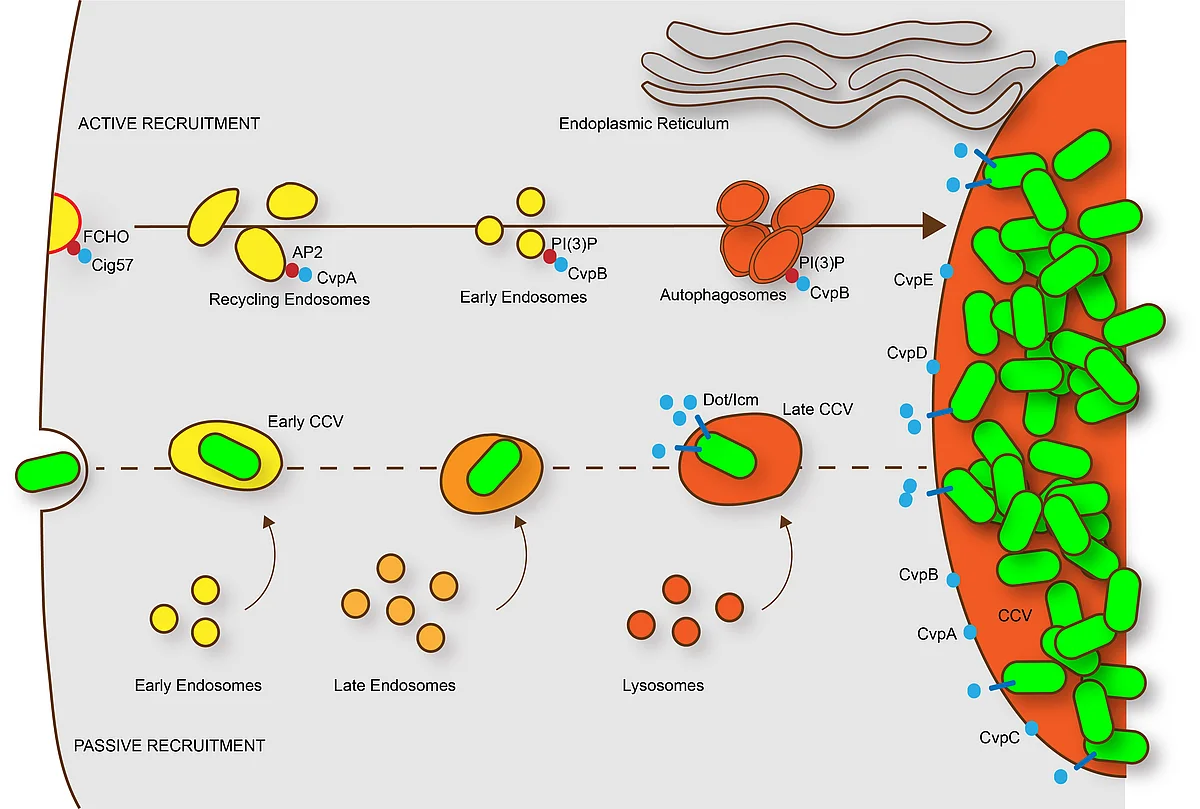
After uptake into mammalian cells by microfilament-dependent endocytosis, C. burnetii remains in a membrane-bound vacuole. This C. burnetii-containing vacuole (CCV) initially appears to mature similarly to phagosomes containing avirulent bacteria, undergoing fusion with endosomes and lysosomes, resulting in the formation of a phagolysosomal compartment. However, C. burnetii delays the maturation of the CCV probably through interactions between the early CCV and the autophagic pathway. How C. burnetii mediates establishment of the phagolysosomal-like compartment in which it resides and replicates, is not well understood. However, a functional type IV secretion system (T4SS) is required, suggesting that bacterial effector proteins may directly influence biogenesis of the C. burnetii-occupied vacuole.
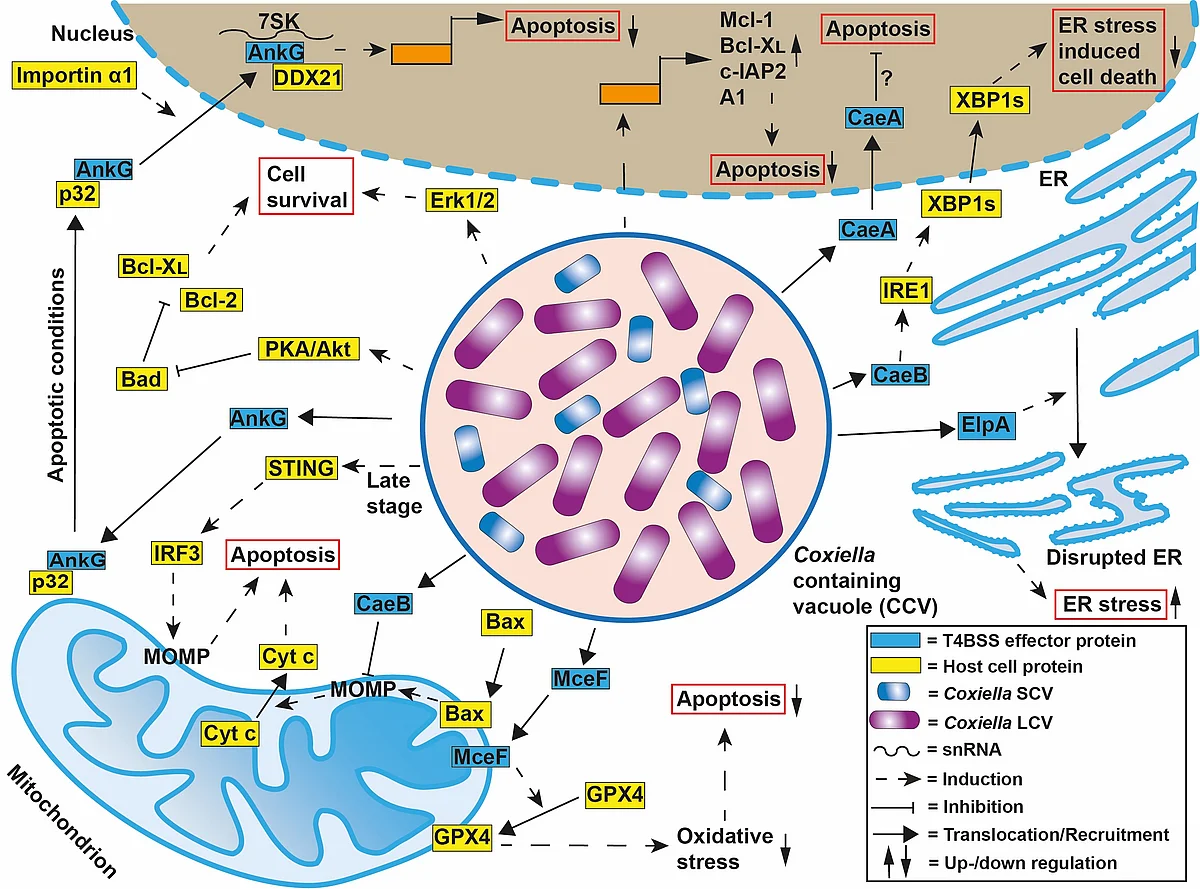
C. burnetii has been shown to inhibit host cell apoptosis. Apoptosis is a programmed cell death pathway that is crucial for immune system maintenance and removal of damaged or infected cells. It was demonstrated that C. burnetii infection inhibits the induction of the intrinsic cell death pathway by preventing cytochrome C release from mitochondria and consequently inhibiting caspase 3 activation. Another study showed that C. burnetii infection inhibited not only the intrinsic but also the extrinsic apoptosis pathway. While the mechanism(s) of C. burnetii-induced inhibition of host cell apoptosis is not well understood, it is clear that it depends on a functional T4SS.
For general information about C. burnetii or Q fever please visit our website: https://q-gaps.de/ or https://q-gaps.de/en/
Our current projects
1.) Analysis of the T4SS effector-induced inhibition of apoptosis
To determine the diverse mechanisms employed by Coxiella burnetii to prevent host cell apoptosis, we will analyze the function of Coxiella burnetii T4SS substrates that interfere with signaling through the intrinsic and extrinsic apoptotic pathways.
2.) Induction and regulation of Coxiella burnetii persistence by microenvironmental factors
We hypothesize that micromilieu factors are decisive for the outcome of C. burnetii infection. Indeed, our results suggest that a micromilieu with reduced oxygen availability (hypoxia) induces C. burnetii persistence. Therefore, we are currently investigating how hypoxia influences the host cell metabolism and immune response during infection and how this triggers bacterial persistence. Furthermore, we are analyzing the functional consequence of C. burnetii persistence for the host-pathogen interaction. (For more information please see: https://www.immunomicrotope.de)
3.) Egress mechanisms of Coxiella burnetii
While we have information about the initial step of C. burnetii uptake and the maturation process of the C. burnetii-containing vacuole, it is largely unknown how C. burnetii spreads, i.e. how it exits its host cell to enter new target cells. Therefore, the aim of this project is to define the mechanism of egress of C. burnetii. (For more information please see: www.spp2225.rwth-aachen.de).

